Модель индукции синтеза IgE
Основная биологическая роль IgE связана с его уникальной способностью связываться с поверхностью тучных клеток и базофилов человека. Только эти клетки несут высокоаффинный гликопротеиновый рецептор (относительная молекулярная масса =130 000 дальтон) для Fc-участка молекулы IgE (FceRl). На поверхности одного базофила имеются 40 000-100 000 рецепторов, которые связывают от 5 000 до 40 000 молекул IgE [3, 7]. На других клетках (В-лимфоцитах, макрофагах, эозинофилах и тромбоцитах) были выявлены низкоаффинные рецепторы для IgE (FceRII). Надмембранная часть этих рецепторов легко поддается протеолитическому отщеплению от клеточной мембраны, превращаясь в растворимый FceRII, идентичный молекуле CD23, обладающий способностью связывать IgE. В связи с этой способностью та же молекула получила название «IgE связывающий фактор>> (IgE-BF). Этот фактор ингибирует связывание IgE не только с FceRII, но и с FceRI, участвуя, таким образом, в регуляции IgE-зависимых аллергических реакций. Кроме того, этот фактор участвует в регуляции продукции IgE: Т-хелперы, несущие FceRII-peцепторы (CD23), продуцируют фактор, усиливающий гликозилирование IgE-BF, а Т-лимфоциты со свойствами супрессоров продуцируют фактор, ингибирующий гликозилирование, гликозилированный IgE-BF потенцирует, а негликозилированный фактор угнетает продукцию IgE. Однако регуляция продукции IgE - это гораздо более сложный процесс (Рисунок 1). Выявлена ассоциация специфического IgE-ответа с определенным HLA-DR гаплотипом.
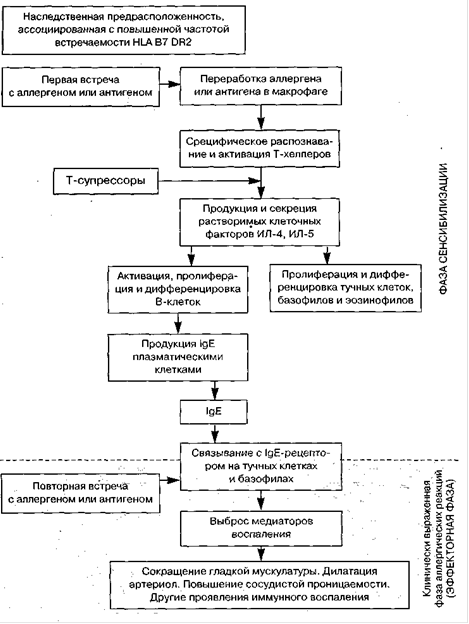
Рисунок 1. Схема продукции IgE, её регуляции и IgE-опосредованных эффектов.
Начальной фазой антиген(аллерген)индуцированного этапа является активация В-лимфоцитов, которая требует двух сигналов: от непосредственно распознающих: антигены Ig-рецепторов В-лимфоцитов и от Т-хелперов. непосредственно и через цитокины, продуцируемые активированными Т-хелперами. Последние являются наиболее важным звеном в регуляции синтеза иммуноглобулинов, так как запускают также этапы пролиферации и дифференцировки активированных В-лимфоцитов в плазматические (антите-лопродуцирующие) клетки. На этом этапе синтез IgE. по сравнению с иммуноглобулинами других классов, имеет некоторые особенности (Рисунок 2). Среди цитокинов, контролирующих продукцию IgE, есть два цитокина, оказывающих разнонаправленное действие на его синтез: интерлей-кин-4 (IL-4) стимулирует, а интерферон-гамма (IFN-gamma) - угнетает. Растворимые низкоаффинные рецепторы FceRII (CD23) в ассоциации с IL-4 способствуют дифференцировке В-лимфоцитов в IgE-синтезирующие клетки, a IFN-gamma ингибирует этот процесс [7-9].
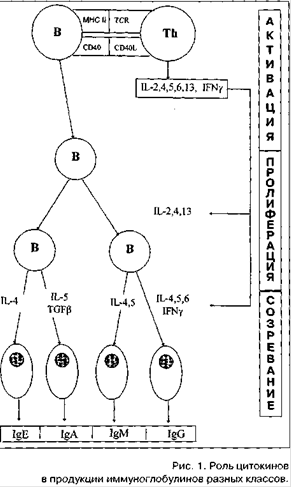
Рисунок 2. Роль цитокинов в продукции иммуноглобулинов разных классов.
Дегрануляция тучных клеток и базофилов наступает, когда две связанные с мембраной клеток молекулы IgE соединяются с антигеном. После этого происходит выброс медиаторов воспаления вследствие активации серинэстеразы, связанной с клеточной мембраной; притока внутрь клетки ионов кальция; образования обладающих способностью к слиянию липидов и последующего соединения периграну-лярной мембраны с плазматической мембраной; снижения уровня внутриклеточного циклического 3',5'-аденозинмонофосфата; сокращения микрофиламентов и агрегация микроканальцев. По мере развития этих процессов гранула с медиатором сначала сообщается с внеклеточной средой при помощи «цистерн», а затем, в конечном итоге, выталкивается из клетки [4,8,9].
Помимо участия в аллергических реакциях I (немедленного) типа, IgE также принимает участие в защитном противогельминтном иммунитете (Рисунок 3), что обусловлено существованием перекрестного связывания между IgE и антигеном гельминтов. Последний, проникая через мембрану слизистой оболочки, "садится" на тучные клетки и вызывает их дегрануляцию. Медиаторы воспаления повышают проницаемость капилляров и слизистых оболочек, в результате чего IgG и лейкоциты выходят из кровотока. К гельминтам, покрытым IgG, присоединяются эозинофилы, которые выбрасывают содержимое своих гранул и таким образом убивают гельминтов [4, 9].